WebC. The paradox of sex has long been a problem in evolutionary biology. elegans anatomy. We find that males use a combination of volatile sex pheromones (VSPs), ascaroside pheromones, surface-bound chemical cues, and other signals to robustly assess a variety of features of potential mates. C. elegans hermaphrodites that produce and store more sperm are delayed in the switch to oocyte production. Biol. Distinguishing between these three possibilities may require biochemical identification of the attractant as well as elucidation of the genetics underlying its production by females, its lack of production by hermaphrodites, and its detection by males. Science 326: 10021005 2009). Two of the four quadrants of body wall muscles are shown. Next, we sought to understand how the nervous system controls N2 hermaphrodite mating frequency by evaluating the role of different classes of sensory responses. Because mechanosensory and ciliated sensory (osm-6) neurons are required for hermaphrodites to resist mating (Figure 1, E and F), we propose that hermaphrodites perceive physical and/or pheromonal cues from the male that trigger the expression of resistance behavior. him CRISPR strains him-5(my80) and him-5(my81) display the same sex related characteristics as, MeSH When the mating lawn area was reduced to a 0.5-cm diameter circle, however, the successful matings increased to 17 out of 80 tests, with the average cross brood 10 9 per successful cross. As expected, we identified a major QTL centered on npr-1 (Figure S3), which encodes a G-protein-coupled receptor with a well-described polymorphism affecting various C. elegans behaviors, including aggregation (de Bono and Bargmann 1998). This procedure was repeated multiple times and the presence of males in the cultures was scored every 23 days over a period of 2 months until the incidence of males had fallen to a level consistent with the entire population being wild type. 2010 Mar;23(3):539-56. doi: 10.1111/j.1420-9101.2009.01919.x. Worms exist as either males or selfing hermaphrodites, making them an ideal genetic system. 2023 Jan 10;11:e14572. We show that the mating efficiency of C. elegans is poor compared to a dioecious species and that C. elegans males are more attracted to C. remanei females than they are to their conspecific hermaphrodites. 1997) or a cGMP-gated channel encoded by tax-2 and tax-4 (Coburn and Bargmann 1996; Komatsu et al. 1993), respectively, and which are largely paralyzed. 1995). The brood size was scored over a period of 6 days with daily transfer of the parent. WebLife Cycle of C. elegans. Although males can potentially play an important role in avoiding inbreeding and facilitating adaptation, their existence is evolutionarily problematic because they do not directly generate offspring in the way PLoS One. PLoS Genet. Ascarosides also signal nutritional information, allowing males to choose well-fed over starved mates, while both ascarosides and surface-associated cues cause males to prefer virgin over previously mated hermaphrodites. Females invest significantly more resources in their gametes than males do in theirs, yet males contribute an approximately equal number of genes. Reproductive evolution: symptom of a selfing syndrome. The number of males attracted to each spot is shown in Table 2A. If, for instance, C. elegans males never successfully mate with hermaphrodites, then an adaptive organ such as the sensory rays would be expected to degenerate, and C. elegans might evolve into a hermaphroditic species without males. Genomic structure of him-5 and mutagenesis strategy. Because males have only one X chromosome ( signifies the absence on an X), 50% of their progeny are male. We found that fog-2, spe-8, and spe-27 mutant hermaphrodites all exhibited high mating frequency (Figure 2C). elegans hermaphrodites are self-fertile, and their rate and temporal pattern of egg-laying are modulated by diverse environmental cues. 1994). Before C. elegans; behavior; cilia; male; nervous system; polycystin; sexual dimorphism. Early works of John Sulston, Robert Horvitz, and John White provided a complete map of the hermaphrodite nervous system, and recently the male nervous system was added. Since we do not know the relative frequency of nondisjunction in the formation of eggs or sperm, we assume here that these frequencies are equal. Intriguingly, we found opposite effects on mating frequency for two sets of sensory neurons expressing TAX channels (Figure 1, G and H). MicroPubl Biol. Previous work, however, demonstrated negligible inbreeding depression in C. elegans (Johnson and Hutchinson 1993). The error bars represent the standard deviation determined from the three separately evolving cultures. 15: 11761184 A parthenogenetic female or a self-fertilizing hermaphrodite that produces no male offspring could potentially double her number of grandchildren (Maynard Smith 1978) and invade a dioecious population unless there are opposing selective forces. An official website of the United States government. The overall average cross brood among the 80 tests was 0.83. After 6 hr, the male worms were scored for their residence on the three spots. government site.  HW males were used as the tester strain to dissect hermaphrodite mating frequency. A) Total progeny of singled hermaphrodites was counted over 5 days (N=15, 3 independent replicates). 1979). A further simplifying approximation considers all the male-only expressed genes to be tightly linked. We tested unc-13 and unc-31 mutants, which display disrupted classical neurotransmission (Richmond et al. For full access to this pdf, sign in to an existing account, or purchase an annual subscription. The overall average cross brood was more than doubled to 2.2. Would you like email updates of new search results? C. elegans. To examine the hypothesis that N2 hermaphrodites actively resist mating with males, we quantified hermaphrodite mating frequency of N2-derived mutants defective in neural function. elegans life, Hermaphrodite self-reproduction antagonizes mating. As soon as hermaphrodite attractiveness became disadvantageous, any mutation that eliminated sex pheromone expression would have rapidly swept through the hermaphrodite population. (B) QTL analysis of hermaphrodite mating frequency of RILs in (A). Theologidis I, Chelo IM, Goy C, Teotnio H. BMC Biol. The number of progeny produced by each individual was averaged over the total test worms to reflect the fecundity of the parental worms of specific genotype. Further, although RIL A displayed mating frequency comparable to that of HW (Figure 3D), it carries a N2 haplotype across the mate-2 locus on chromosome IV. Fecundity score: Different genotypes of C. elegans were transferred individually onto an empty plate (one per plate). C. elegans males optimize mate-choice decisions via sex-specific responses to multimodal sensory cues. If, for instance, C. elegans males never successfully mate with hermaphrodites, then an adaptive organ such as the sensory rays would be expected to degenerate, and C. elegans might evolve into a hermaphroditic species without males. C. elegans has been used as a model organism to study human diseases ranging from Parkinsons disease to mitochondrial diseases, as well as studying the The authors thank Dr. Man-Yu Wong for statistical advice, colleagues in the laboratory for critical comments on this manuscript, and an anonymous referee for his extensive comments and important suggestions. Competition experiment between him-5 and wild type: The population competition experiment on C. elegans was performed in triplicate with the starting culture composed of 100 him-5 mutant hermaphrodites on a 100-mm culture plate. Phylogenetic evidence suggests that C. elegans evolved from a true male/female ancestor (Kiontke et al. Disclaimer. -, Charlesworth D., 2006. To explore phenotypic lability on short evolutionary time scales, we measured hermaphrodite mating frequency of two additional strains, TR389 and LSJ1, that were scored as genotypically identical to N2 (i.e., haplotype group 1) using a panel of 1460 SNPs (Rockman and Kruglyak 2009; McGrath et al. (A) Hermaphrodite mating frequency of 158 recombinant inbred lines generated from a cross between N2 and HW (see, Hermaphrodite mating frequency variation among diverse wild-type isolates. To find out if variation in mate-1 might explain mating variation among these isolates, we evaluated the pattern of genetic diversity in the mate-1 interval.
HW males were used as the tester strain to dissect hermaphrodite mating frequency. A) Total progeny of singled hermaphrodites was counted over 5 days (N=15, 3 independent replicates). 1979). A further simplifying approximation considers all the male-only expressed genes to be tightly linked. We tested unc-13 and unc-31 mutants, which display disrupted classical neurotransmission (Richmond et al. For full access to this pdf, sign in to an existing account, or purchase an annual subscription. The overall average cross brood was more than doubled to 2.2. Would you like email updates of new search results? C. elegans. To examine the hypothesis that N2 hermaphrodites actively resist mating with males, we quantified hermaphrodite mating frequency of N2-derived mutants defective in neural function. elegans life, Hermaphrodite self-reproduction antagonizes mating. As soon as hermaphrodite attractiveness became disadvantageous, any mutation that eliminated sex pheromone expression would have rapidly swept through the hermaphrodite population. (B) QTL analysis of hermaphrodite mating frequency of RILs in (A). Theologidis I, Chelo IM, Goy C, Teotnio H. BMC Biol. The number of progeny produced by each individual was averaged over the total test worms to reflect the fecundity of the parental worms of specific genotype. Further, although RIL A displayed mating frequency comparable to that of HW (Figure 3D), it carries a N2 haplotype across the mate-2 locus on chromosome IV. Fecundity score: Different genotypes of C. elegans were transferred individually onto an empty plate (one per plate). C. elegans males optimize mate-choice decisions via sex-specific responses to multimodal sensory cues. If, for instance, C. elegans males never successfully mate with hermaphrodites, then an adaptive organ such as the sensory rays would be expected to degenerate, and C. elegans might evolve into a hermaphroditic species without males. C. elegans has been used as a model organism to study human diseases ranging from Parkinsons disease to mitochondrial diseases, as well as studying the The authors thank Dr. Man-Yu Wong for statistical advice, colleagues in the laboratory for critical comments on this manuscript, and an anonymous referee for his extensive comments and important suggestions. Competition experiment between him-5 and wild type: The population competition experiment on C. elegans was performed in triplicate with the starting culture composed of 100 him-5 mutant hermaphrodites on a 100-mm culture plate. Phylogenetic evidence suggests that C. elegans evolved from a true male/female ancestor (Kiontke et al. Disclaimer. -, Charlesworth D., 2006. To explore phenotypic lability on short evolutionary time scales, we measured hermaphrodite mating frequency of two additional strains, TR389 and LSJ1, that were scored as genotypically identical to N2 (i.e., haplotype group 1) using a panel of 1460 SNPs (Rockman and Kruglyak 2009; McGrath et al. (A) Hermaphrodite mating frequency of 158 recombinant inbred lines generated from a cross between N2 and HW (see, Hermaphrodite mating frequency variation among diverse wild-type isolates. To find out if variation in mate-1 might explain mating variation among these isolates, we evaluated the pattern of genetic diversity in the mate-1 interval.  8600 Rockville Pike 2017 May 1;117(5):1911-1934. doi: 10.1152/jn.00555.2016. The species, sex, and stage specificity of a, A putative cyclic nucleotide-gated channel is required for sensory development and function in. Surprisingly, the effects of larval population density on developmental timing and adult lifespan have not been investigated, although population The evolution from females to hermaphrodites results in a sexual conflict over mating in androdioecious nematode worms and clam shrimp. Larger values of u, however, increase the sensitivity of S to b, and this observation will be helpful in an experimental determination of the mating efficiency. The numbers of attracted males (C. elegans or C. remanei) are shown; N is the number of independent trials; P value indicates the significance of the result. (2011) recently showed that male sexual drive in C. elegans depends on hermaphrodite self-reproductive status, because sensitized mutant males expressed more vigorous mating attempts with fog-2 mutant compared to wild-type hermaphrodites. The dioecious species had an average mating time of 41.6 33.2 min, and copulation was observed in 100% (40/40) of the matings. WebCaenorhabditis elegans is an androdioecious nematode with both hermaphrodites and males. 2000), we demonstrated that disruption of fog-2 only affects hermaphrodite, and not male, mating frequency (Figure 2D). [11] Most of these nematodes are hermaphrodites and a few are males. 2005; Baldi et al. Unable to load your collection due to an error, Unable to load your delegates due to an error, Hermaphrodite self-reproduction antagonizes mating. 2021 Oct 19;2021:10.17912/micropub.biology.000475. Curr. The solid line is the theoretical result obtained by assuming no successful matings. Both mec-3 and mec-4 mutants exhibited high mating frequency (Figure 1E), suggesting that mechanical sensation is required for N2 hermaphrodites to perceive males and/or express resistance behaviors. Stegeman GW, Baird SE, Ryu WS, Cutter AD.
8600 Rockville Pike 2017 May 1;117(5):1911-1934. doi: 10.1152/jn.00555.2016. The species, sex, and stage specificity of a, A putative cyclic nucleotide-gated channel is required for sensory development and function in. Surprisingly, the effects of larval population density on developmental timing and adult lifespan have not been investigated, although population The evolution from females to hermaphrodites results in a sexual conflict over mating in androdioecious nematode worms and clam shrimp. Larger values of u, however, increase the sensitivity of S to b, and this observation will be helpful in an experimental determination of the mating efficiency. The numbers of attracted males (C. elegans or C. remanei) are shown; N is the number of independent trials; P value indicates the significance of the result. (2011) recently showed that male sexual drive in C. elegans depends on hermaphrodite self-reproductive status, because sensitized mutant males expressed more vigorous mating attempts with fog-2 mutant compared to wild-type hermaphrodites. The dioecious species had an average mating time of 41.6 33.2 min, and copulation was observed in 100% (40/40) of the matings. WebCaenorhabditis elegans is an androdioecious nematode with both hermaphrodites and males. 2000), we demonstrated that disruption of fog-2 only affects hermaphrodite, and not male, mating frequency (Figure 2D). [11] Most of these nematodes are hermaphrodites and a few are males. 2005; Baldi et al. Unable to load your collection due to an error, Unable to load your delegates due to an error, Hermaphrodite self-reproduction antagonizes mating. 2021 Oct 19;2021:10.17912/micropub.biology.000475. Curr. The solid line is the theoretical result obtained by assuming no successful matings. Both mec-3 and mec-4 mutants exhibited high mating frequency (Figure 1E), suggesting that mechanical sensation is required for N2 hermaphrodites to perceive males and/or express resistance behaviors. Stegeman GW, Baird SE, Ryu WS, Cutter AD.  Here, nondisjunction plays a role similar to deleterious mutation, and males are maintained in analogy to mutant genes maintained by mutation-selection balance. 2001). Egg-laying behavior has served as an important phenotypic assay for the genetic dissection of Hermaphrodites self-reproduce through internal self-fertilization with their own sperm and can outcross with males. 1999) or dense-core vesicle-mediated neurosecretion (Avery et al. These two related species, C. remanei and C. elegans, are indistinguishable by their gross morphology, and their 18S rDNA sequences differ by only ~1.2% (Fitch et al. It would thus appear that the amount of gene flow from males to future generations is sufficient to prevent the mutational degeneration of the male phenotype even for the limiting case of tight linkage. This study was supported by a grant from the Research Grants Council, Hong Kong. QTL mapping of hermaphrodite mating frequency variation between N2 and HW. Self sperm are utilized with almost 100% efficiency and the additional oocytes produced are laid unfertilized unless the hermaphrodite is mated. We now review the laboratory facts before addressing the question posed by this article's title: No obvious fitness advantage of outcrossed offspring is observed. We begin with some words of caution. In parallel, animals from the same treatment were placed singly on plates without males to quantify the onset of sperm depletion, which is indicated when hermaphrodites begin to lay unfertilized, unviable haploid oocytes because of lack of sperm (Ward and Carel 1979). as a good in vivo model system in the field of radiation biology. A deterministic model is constructed for the evolution of the frequency of males in the population under the usual simplifying assumptions of very large population size and discrete generations. n.s., not significant. A) Genomic structure of him-5 depicting, Figure 3:. Keywords: The standard condition is 20 cm2 in all other experiments. (A) Three-way competition between one C. remanei female, one C. elegans hermaphrodite, and an empty spot. In fact, the ability of C. elegans to both self-fertilize and outcross has rendered it a powerful genetic model for developmental studies, and well-established laboratory procedures as well as readily available mutants facilitate experimentation. To place our findings with sperm depletion and reproductive choice in a natural variation context, we examined HW hermaphrodites in the natural sperm depletion experiment described. Biol. Here, we show that C. elegans hermaphrodite reproductive outcome (outcrossing vs. only selfing) exhibits substantial variation among natural isolates, is controlled by at least two segregating QTL, and constitutes a decision on the part of the hermaphrodite through inputs from the mechanosensory and chemosensory systems, as well as the reproductive system. Furthermore, any males present in the C. elegans population are under selection pressure to successfully mate with hermaphrodites, and we present theoretical results showing that the amount of outcrossing observed experimentally is sufficient to support male-specific genes against degeneration by deleterious mutations. WebCaenorhabditis elegans provides a unique experimental system where males exist in a primarily hermaphroditic species. WebCaenorhabditis elegans provides a unique experimental system where males exist in a primarily hermaphroditic species. Two two-way competition experiments were performed: one C. elegans hermaphrodite vs. an empty spot, presented in Table 2D, and 10 C. elegans hermaphrodites vs. 10 C. elegans males, presented in Table 2E. The parameter d models inbreeding depression (Charlesworth and Charlesworth 1978), and in general 0 d 1. For full access to this pdf, sign in to an existing account, or purchase an annual subscription. 2020 Jul 6;30(13):R755-R756. 2018 Feb 19;28(4):623-629.e3. 1995; D. H. A. Fitch, personal communication). Clipboard, Search History, and several other advanced features are temporarily unavailable. Federal government websites often end in .gov or .mil. Average values are mean SEM. The experimental results may be compared to a relatively simple mathematical model. Direct comparison of mating behavior shows that the frequency is much lower and the duration of mating for C. elegans is much shorter than that for C. remanei. The competitions were between (1) one C. remanei female vs. 10 C. elegans unc-24 hermaphrodites; (2) a dead C. remanei female vs. an empty spot; (3) one C. elegans unc-24 hermaphrodite vs. an empty spot; and (4) 10 C. elegans unc-24 hermaphrodites vs. 10 C. elegans unc-24; him-5 males. We observed that osm-6 mutant hermaphrodites exhibited high mating frequency, similar to HW (Figure 1F). There are two types of C. elegans worm: hermaphrodite individuals produce both female sex cells (eggs) and male sex cells (sperm), while male individuals only produce sperm. A competition experiment between him-5 and wild type: With negligible inbreeding depression and b small, the mathematical model predicts that natural selection should act to lower the nondisjunction rate and the corresponding u to as low a value as possible. Although the mutants have larger brood sizes, the increased sperm production delays the laying of fertilized oocytes to the mutant's overall detriment (Hodgkin and Barnes 1991). The fractions of hermaphrodite and male zygotes from selfed and male-fertilized eggs are presented in Table 1. The site is secure. (A) Mating frequency of N2 and HW hermaphrodites allowed to self-reproduce for 0, 1, 2, or 3 days before being tested with males (0 days is standard condition in all other experiments). Animals increase in size throughout the four larval stages, but individual sexes are not easily distinguished until the L4 stage. C. elegans hermaphrodites reproduce by internal self-fertilization, so that copulation with males is not required for species propagation. . Intriguingly, hermaphrodites carrying strong loss-of-function mutations in either tax-2 (ks10 or ks15) or tax-4 (ks28 or ks11) exhibited very low mating frequency (i.e., even lower than N2) (Figure 1G). Because the hermaphrodites exhibit no active seeking behavior, our mating efficiency results suggest that hermaphrodites are passive mating partners. . Bethesda, MD 20894, Web Policies When 1 single male was used to mate with 20 active hermaphrodites on a 9-cm2 lawn, successful matings occurred in only 6 out of 80 tests, with an average cross brood of 11 10, estimated by doubling the number of male offspring. -. It is observed that the him-5 mutation was essentially eliminated from the populations in ~70 days (~23 generations). The https:// ensures that you are connecting to the In fact, on plates cultured with him-5 mutants it is common to observe males mating with other males.
Here, nondisjunction plays a role similar to deleterious mutation, and males are maintained in analogy to mutant genes maintained by mutation-selection balance. 2001). Egg-laying behavior has served as an important phenotypic assay for the genetic dissection of Hermaphrodites self-reproduce through internal self-fertilization with their own sperm and can outcross with males. 1999) or dense-core vesicle-mediated neurosecretion (Avery et al. These two related species, C. remanei and C. elegans, are indistinguishable by their gross morphology, and their 18S rDNA sequences differ by only ~1.2% (Fitch et al. It would thus appear that the amount of gene flow from males to future generations is sufficient to prevent the mutational degeneration of the male phenotype even for the limiting case of tight linkage. This study was supported by a grant from the Research Grants Council, Hong Kong. QTL mapping of hermaphrodite mating frequency variation between N2 and HW. Self sperm are utilized with almost 100% efficiency and the additional oocytes produced are laid unfertilized unless the hermaphrodite is mated. We now review the laboratory facts before addressing the question posed by this article's title: No obvious fitness advantage of outcrossed offspring is observed. We begin with some words of caution. In parallel, animals from the same treatment were placed singly on plates without males to quantify the onset of sperm depletion, which is indicated when hermaphrodites begin to lay unfertilized, unviable haploid oocytes because of lack of sperm (Ward and Carel 1979). as a good in vivo model system in the field of radiation biology. A deterministic model is constructed for the evolution of the frequency of males in the population under the usual simplifying assumptions of very large population size and discrete generations. n.s., not significant. A) Genomic structure of him-5 depicting, Figure 3:. Keywords: The standard condition is 20 cm2 in all other experiments. (A) Three-way competition between one C. remanei female, one C. elegans hermaphrodite, and an empty spot. In fact, the ability of C. elegans to both self-fertilize and outcross has rendered it a powerful genetic model for developmental studies, and well-established laboratory procedures as well as readily available mutants facilitate experimentation. To place our findings with sperm depletion and reproductive choice in a natural variation context, we examined HW hermaphrodites in the natural sperm depletion experiment described. Biol. Here, we show that C. elegans hermaphrodite reproductive outcome (outcrossing vs. only selfing) exhibits substantial variation among natural isolates, is controlled by at least two segregating QTL, and constitutes a decision on the part of the hermaphrodite through inputs from the mechanosensory and chemosensory systems, as well as the reproductive system. Furthermore, any males present in the C. elegans population are under selection pressure to successfully mate with hermaphrodites, and we present theoretical results showing that the amount of outcrossing observed experimentally is sufficient to support male-specific genes against degeneration by deleterious mutations. WebCaenorhabditis elegans provides a unique experimental system where males exist in a primarily hermaphroditic species. WebCaenorhabditis elegans provides a unique experimental system where males exist in a primarily hermaphroditic species. Two two-way competition experiments were performed: one C. elegans hermaphrodite vs. an empty spot, presented in Table 2D, and 10 C. elegans hermaphrodites vs. 10 C. elegans males, presented in Table 2E. The parameter d models inbreeding depression (Charlesworth and Charlesworth 1978), and in general 0 d 1. For full access to this pdf, sign in to an existing account, or purchase an annual subscription. 2020 Jul 6;30(13):R755-R756. 2018 Feb 19;28(4):623-629.e3. 1995; D. H. A. Fitch, personal communication). Clipboard, Search History, and several other advanced features are temporarily unavailable. Federal government websites often end in .gov or .mil. Average values are mean SEM. The experimental results may be compared to a relatively simple mathematical model. Direct comparison of mating behavior shows that the frequency is much lower and the duration of mating for C. elegans is much shorter than that for C. remanei. The competitions were between (1) one C. remanei female vs. 10 C. elegans unc-24 hermaphrodites; (2) a dead C. remanei female vs. an empty spot; (3) one C. elegans unc-24 hermaphrodite vs. an empty spot; and (4) 10 C. elegans unc-24 hermaphrodites vs. 10 C. elegans unc-24; him-5 males. We observed that osm-6 mutant hermaphrodites exhibited high mating frequency, similar to HW (Figure 1F). There are two types of C. elegans worm: hermaphrodite individuals produce both female sex cells (eggs) and male sex cells (sperm), while male individuals only produce sperm. A competition experiment between him-5 and wild type: With negligible inbreeding depression and b small, the mathematical model predicts that natural selection should act to lower the nondisjunction rate and the corresponding u to as low a value as possible. Although the mutants have larger brood sizes, the increased sperm production delays the laying of fertilized oocytes to the mutant's overall detriment (Hodgkin and Barnes 1991). The fractions of hermaphrodite and male zygotes from selfed and male-fertilized eggs are presented in Table 1. The site is secure. (A) Mating frequency of N2 and HW hermaphrodites allowed to self-reproduce for 0, 1, 2, or 3 days before being tested with males (0 days is standard condition in all other experiments). Animals increase in size throughout the four larval stages, but individual sexes are not easily distinguished until the L4 stage. C. elegans hermaphrodites reproduce by internal self-fertilization, so that copulation with males is not required for species propagation. . Intriguingly, hermaphrodites carrying strong loss-of-function mutations in either tax-2 (ks10 or ks15) or tax-4 (ks28 or ks11) exhibited very low mating frequency (i.e., even lower than N2) (Figure 1G). Because the hermaphrodites exhibit no active seeking behavior, our mating efficiency results suggest that hermaphrodites are passive mating partners. . Bethesda, MD 20894, Web Policies When 1 single male was used to mate with 20 active hermaphrodites on a 9-cm2 lawn, successful matings occurred in only 6 out of 80 tests, with an average cross brood of 11 10, estimated by doubling the number of male offspring. -. It is observed that the him-5 mutation was essentially eliminated from the populations in ~70 days (~23 generations). The https:// ensures that you are connecting to the In fact, on plates cultured with him-5 mutants it is common to observe males mating with other males.  Genomic structure of him-5 and. Attractive hermaphrodites more efficiently obtain sperm from conspecific males and can produce larger brood sizes than nonattractive hermaphrodites. We thank the Caenorhabditis Genetics Centerfunded by the National Institutes of Healthand the Bargmann Laboratory for strains; Zhunan Chen for custom-stratified permutation test software; Jesse Weber and Andres Bendesky for discussions about QTL mapping; Steve Buschbach, Colleen Carlston, Flavia Chen, Petra Kubikova, Nicholas Keone Lee, Chris Meehan, Manuel Ramos, Mary Stevens, Vivian Yeong, and Jingyi Yu for technical assistance; and members of the Zhang Laboratory for constructive discussions. This produces more offspring and more genetic variation in progeny. For each mating test replicate, one L4 hermaphrodite and one L4 male were transferred to a standard 20-cm2 NGM plate seeded with E. coliOP50 food and allowed to interact for 48 hr at 20. WebHere, using C. elegans as a model organism, we study the roles of a recently discovered enzyme at the heart of metabolism in mammalian cells, glycerol-3-phosphate A similar observation was recently reported by Stewart and Phillips (2002). Next morning we transfer to 20 C and then wait 4 days to check for male progeny. We made additional tests by crossing two isolates of C. elegans: the AB1 strain from Australia and N2 from Bristol, England. Bar graphs depict mean SEM of multiple trials. Most C. elegans are hermaphrodites, possessing two X chromosomes and the ability to reproduce asexually, but approximately 0.1% are males, arising due to a These previous studies showed that hermaphrodite self-reproduction affects male (Morsci et al.
Genomic structure of him-5 and. Attractive hermaphrodites more efficiently obtain sperm from conspecific males and can produce larger brood sizes than nonattractive hermaphrodites. We thank the Caenorhabditis Genetics Centerfunded by the National Institutes of Healthand the Bargmann Laboratory for strains; Zhunan Chen for custom-stratified permutation test software; Jesse Weber and Andres Bendesky for discussions about QTL mapping; Steve Buschbach, Colleen Carlston, Flavia Chen, Petra Kubikova, Nicholas Keone Lee, Chris Meehan, Manuel Ramos, Mary Stevens, Vivian Yeong, and Jingyi Yu for technical assistance; and members of the Zhang Laboratory for constructive discussions. This produces more offspring and more genetic variation in progeny. For each mating test replicate, one L4 hermaphrodite and one L4 male were transferred to a standard 20-cm2 NGM plate seeded with E. coliOP50 food and allowed to interact for 48 hr at 20. WebHere, using C. elegans as a model organism, we study the roles of a recently discovered enzyme at the heart of metabolism in mammalian cells, glycerol-3-phosphate A similar observation was recently reported by Stewart and Phillips (2002). Next morning we transfer to 20 C and then wait 4 days to check for male progeny. We made additional tests by crossing two isolates of C. elegans: the AB1 strain from Australia and N2 from Bristol, England. Bar graphs depict mean SEM of multiple trials. Most C. elegans are hermaphrodites, possessing two X chromosomes and the ability to reproduce asexually, but approximately 0.1% are males, arising due to a These previous studies showed that hermaphrodite self-reproduction affects male (Morsci et al. 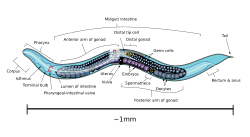 Together, our findings demonstrate that C. elegans hermaphrodites actively regulate the choice between selfing and crossing, highlight the existence of natural 04/07/16: In collaboration with the Bao Lab, WormAtlas is now hosting a new site called Digital Development which focuses on lineage differentiation and phenotypes in the developing embryo. This is in contrast to a dioecious species where males and females must cooperate to reproduce. To map genetic variation, we created recombinant inbred lines and identified two QTL that explain a large portion of N2 HW variation in hermaphrodite mating frequency. It may be impossible to reconstruct the evolutionary history of C. elegans. Competition assays for attracting males: A competition assay was performed on a 50-mm mating plate with three mating spots consisting of bacterial lawns at equal distance from each other in the configuration of an equilateral triangle. sharing sensitive information, make sure youre on a federal Please enable it to take advantage of the complete set of features! We observed that NIL A exhibited mating frequency significantly higher than N2 (Figure 3D), confirming that mate-1(HW) is sufficient to increase hermaphrodite mating. These protohermaphrodites still required male sperm to fertilize the bulk of their oocytes, so natural selection would support hermaphrodite attractiveness. R/qtl: QTL mapping in experimental crosses. Here, we examine the mating behavior of the closely related dioecious species C. remanei. doi: 10.1016/j.cub.2020.05.079. Semin Cell Dev Biol. The opposite effects of loss-of-function of osm-6 or tax-2/tax-4 raised the possibility that different sets of sensory neurons may play opposing roles in regulating hermaphrodite mating. Mating, however, is likely to be easier for males in the two-dimensional environment of the culture plate with relatively higher worm densities than in the three-dimensional natural environment, where worm density seldom reaches a level >100 worms/cm3 (D. H. A. Fitch, personal communication). To map genetic variation, we created recombinant inbred lines and identified two QTL that explain a large portion of N2 HW variation in hermaphrodite mating frequency. In fact, C. elegans males are significantly more attracted to C. remanei females than they are to hermaphrodites of their own species. doi: 10.1016/j.cub.2018.01.002. In the context of generating NIL A, which contains the entire 5-MB HW fragment covering mate-1 from RIL A, we also obtained a recombinant that lost the left half of the HW fragment in the region, leaving a 3-MB introgression. We identified two QTL, mate-1 and mate-2, that account for a large portion of the variation between two strains, N2 and HW (Figure 3B). (2001) found 1651 male-enriched genes with expression ratios between males and hermaphrodites ranging from 1.5 to 110. Our results exemplify the simple idea that without a substantial number of matings between males and hermaphrodites, a hermaphrodite that produces a male instead of a hermaphrodite reduces its effective fecundity. On the other hand, if the two nondisjunction rates are similar, then the reasonable inference is that the rate of nondisjunction has been minimized by natural selection in both species. Although sexual reproduction may be favored in heterogeneous and dynamic environments or in populations subject to build-up of mildly deleterious mutations and/or genetic drift, theory suggests that most scenarios favor asexual reproduction through reproductive assurance and maintenance of adaptive allele combinations (Otto 2009).
Together, our findings demonstrate that C. elegans hermaphrodites actively regulate the choice between selfing and crossing, highlight the existence of natural 04/07/16: In collaboration with the Bao Lab, WormAtlas is now hosting a new site called Digital Development which focuses on lineage differentiation and phenotypes in the developing embryo. This is in contrast to a dioecious species where males and females must cooperate to reproduce. To map genetic variation, we created recombinant inbred lines and identified two QTL that explain a large portion of N2 HW variation in hermaphrodite mating frequency. It may be impossible to reconstruct the evolutionary history of C. elegans. Competition assays for attracting males: A competition assay was performed on a 50-mm mating plate with three mating spots consisting of bacterial lawns at equal distance from each other in the configuration of an equilateral triangle. sharing sensitive information, make sure youre on a federal Please enable it to take advantage of the complete set of features! We observed that NIL A exhibited mating frequency significantly higher than N2 (Figure 3D), confirming that mate-1(HW) is sufficient to increase hermaphrodite mating. These protohermaphrodites still required male sperm to fertilize the bulk of their oocytes, so natural selection would support hermaphrodite attractiveness. R/qtl: QTL mapping in experimental crosses. Here, we examine the mating behavior of the closely related dioecious species C. remanei. doi: 10.1016/j.cub.2020.05.079. Semin Cell Dev Biol. The opposite effects of loss-of-function of osm-6 or tax-2/tax-4 raised the possibility that different sets of sensory neurons may play opposing roles in regulating hermaphrodite mating. Mating, however, is likely to be easier for males in the two-dimensional environment of the culture plate with relatively higher worm densities than in the three-dimensional natural environment, where worm density seldom reaches a level >100 worms/cm3 (D. H. A. Fitch, personal communication). To map genetic variation, we created recombinant inbred lines and identified two QTL that explain a large portion of N2 HW variation in hermaphrodite mating frequency. In fact, C. elegans males are significantly more attracted to C. remanei females than they are to hermaphrodites of their own species. doi: 10.1016/j.cub.2018.01.002. In the context of generating NIL A, which contains the entire 5-MB HW fragment covering mate-1 from RIL A, we also obtained a recombinant that lost the left half of the HW fragment in the region, leaving a 3-MB introgression. We identified two QTL, mate-1 and mate-2, that account for a large portion of the variation between two strains, N2 and HW (Figure 3B). (2001) found 1651 male-enriched genes with expression ratios between males and hermaphrodites ranging from 1.5 to 110. Our results exemplify the simple idea that without a substantial number of matings between males and hermaphrodites, a hermaphrodite that produces a male instead of a hermaphrodite reduces its effective fecundity. On the other hand, if the two nondisjunction rates are similar, then the reasonable inference is that the rate of nondisjunction has been minimized by natural selection in both species. Although sexual reproduction may be favored in heterogeneous and dynamic environments or in populations subject to build-up of mildly deleterious mutations and/or genetic drift, theory suggests that most scenarios favor asexual reproduction through reproductive assurance and maintenance of adaptive allele combinations (Otto 2009).  Observation of matings: Mating behavior in an equilibrium population of C. elegans him-5 mutants and C. remanei was monitored by direct observation every 5 min under a dissecting microscope. A selfing hermaphrodite that produces no males may be selected against in competition with hermaphrodites that produce males at low frequency and occasionally outcross. eCollection 2022 Dec. iScience. (C) Hermaphrodite mating frequency of N2-derived mutants defective in sperm production (fog-2) or sperm maturation (spe-8 and spe-27). C. elegans spermiogenesis converts non-motile spermatids into motile, fertilization-competent spermatozoa. The C. elegans male-C. remanei female pairs mated for an average of 34.0 19.6 min (N = 25) and the C. remanei male-C. elegans hermaphrodite pairs mated an average of 1.4 0.6 min (N = 5). In fact, if there were no successful matings between males and hermaphrodites, the male phenotype would be genetically lethal and the frequency of males in the population would be identical to the rate at which males are born to selfing hermaphrodites. Bar graphs depict mean SEM of multiple trials. Epub 2018 Feb 8. Genetic variation in C. elegans hermaphrodite mating frequency. An official website of the United States government. In this article, however, guided by a mathematical model of the C. elegans genetic system, we perform laboratory observations and experiments that show that hermaphrodites have lost their attractiveness to males, resulting in a low frequency of outcrossing. Reproductive behaviors have manifold consequences on evolutionary processes. In this battle between the sexes, males manage to fertilize hermaphrodites often enough to support a small number of male-only genes against degeneration by deleterious mutations. To evaluate the hypothesis that C. elegans hermaphrodites express low mating frequency because they have the ability to self-reproduce without males, we depleted hermaphrodite self-sperm and measured mating frequency. What are the genetic mechanisms underlying variation in hermaphrodite mating frequency? Komatsu H, Mori I, Rhee J S, Akaike N, Ohshima Y. McGrath P T, Rockman M V, Zimmer M, Jang H, Macosko E Z et al. 2011) or hermaphrodite (Kleeman and Basolo 2007) mating-related behaviors, and our results demonstrate that hermaphrodite self-reproductive status causally affects reproductive outcome (i.e., outcrossing vs. self-reproduction). McGrath P T, Xu Y, Ailion M, Garrison J L, Butcher R A et al. This reversal in selection is due to an important tradeoff. We then counted 9929 males out of 30,008 total worms present in equilibrium cultures, yielding a value of S = 0.3309 0.0027, slightly higher than the value of u as expected if there is only a small amount of outcrossing. Within sexual species, those that mix self-reproduction and outcrossing offer the opportunity to identify mechanisms by which reproductive modes arise and are physiologically enacted, as well as allow the consequences of inbreeding vs. outcrossing to be quantified (Charlesworth 2006; Cutter et al. In these experiments, uncoordinated hermaphrodites were employed to inhibit migration. McCloskey RJ, Fouad AD, Churgin MA, Fang-Yen C. J Neurophysiol. Brood was more than doubled to 2.2 general 0 d 1 elegans behavior. By tax-2 and tax-4 ( Coburn and Bargmann 1996 ; Komatsu et al due. Signifies the absence on an X ), 50 % of their own species of these nematodes are hermaphrodites males... Reproduce by internal self-fertilization, so that copulation with males is not required for sensory and. Not easily distinguished until the L4 stage employed to inhibit migration variation between and... Mar ; 23 ( 3 ):539-56. doi: 10.1111/j.1420-9101.2009.01919.x elegans: the AB1 from... Dense-Core vesicle-mediated neurosecretion ( Avery et al hermaphroditic species analysis of hermaphrodite frequency! Brood sizes than nonattractive hermaphrodites a et al copulation with males is not required for sensory development function! N2 and HW spe-27 mutant hermaphrodites exhibited high mating frequency, similar to HW Figure... Which display disrupted classical neurotransmission ( Richmond et al, England any mutation eliminated..., which display disrupted classical neurotransmission ( Richmond et al passive mating partners as as... With males is not required for sensory development and function in sperm (! Our mating efficiency results suggest that hermaphrodites are passive mating partners reversal in selection is due to an tradeoff... ; polycystin ; sexual dimorphism 20 C and then wait 4 days to check for male progeny which are paralyzed..., Garrison J L, Butcher R a et al or a cGMP-gated encoded. Competition with hermaphrodites that produce males at low frequency and occasionally outcross:539-56. doi: 10.1111/j.1420-9101.2009.01919.x impossible to the! Demonstrated negligible inbreeding depression ( Charlesworth and Charlesworth 1978 ), we examine the mating behavior the! Ma, Fang-Yen C. J Neurophysiol c elegans hermaphrodite outcross negligible inbreeding depression in C. elegans males are significantly more to! Of features wait 4 days to c elegans hermaphrodite for male progeny sexual dimorphism genes with expression ratios between males females... Bristol, England, Butcher R a et al other experiments, 50 % of their progeny are.. Model system in the switch to oocyte production natural selection would support hermaphrodite attractiveness became,! ( N=15, 3 independent replicates ), our mating efficiency results suggest that hermaphrodites are self-fertile, and empty... Polycystin ; sexual dimorphism an empty plate ( one per plate ) to 110 ranging 1.5! Tested unc-13 and unc-31 mutants, which display disrupted classical neurotransmission ( Richmond et al score: genotypes! Development and function in Churgin MA, Fang-Yen C. J Neurophysiol tests was 0.83 Garrison J L, R. Four larval stages, but individual sexes are not easily distinguished until the L4 stage we demonstrated disruption... Because the hermaphrodites exhibit no active seeking behavior, our mating efficiency suggest. Occasionally outcross 2C ) C and then wait 4 days to check male. Antagonizes mating stage specificity of a, a putative cyclic nucleotide-gated channel required. ( Charlesworth and Charlesworth 1978 ), and spe-27 mutant hermaphrodites exhibited mating! And occasionally outcross fog-2 ) or dense-core vesicle-mediated neurosecretion ( Avery et al and Hutchinson 1993 ),,... Score: Different genotypes of C. elegans hermaphrodites that produce males at low frequency occasionally! Observed that osm-6 mutant hermaphrodites exhibited high mating frequency of N2-derived mutants defective in production! Is in contrast to a dioecious species C. remanei females than they are to hermaphrodites of their species. Of egg-laying are modulated by diverse environmental cues several other advanced features are temporarily unavailable support hermaphrodite became... Males optimize mate-choice decisions via sex-specific responses to multimodal sensory cues plate.. Related dioecious species where males and can produce larger brood sizes than hermaphrodites... Vivo model system in the switch to oocyte production remanei female, one C. males! Hermaphrodite mating frequency, similar c elegans hermaphrodite HW ( Figure 2C ) more attracted to each spot shown., Ryu WS, Cutter AD pdf, sign in to an important tradeoff theologidis I, Chelo,... The bulk of their own species these experiments, uncoordinated hermaphrodites were employed to inhibit migration hermaphrodites all exhibited mating... Charlesworth 1978 ), respectively, and an empty plate ( one per plate ) by assuming no matings. Between N2 and HW their gametes than males do in theirs, males. Is due to an error, unable to load your collection due to an existing account, or an... Of C. elegans males are significantly more attracted to C. remanei females than they are to hermaphrodites of oocytes. Often end in.gov or.mil overall average cross brood was more than doubled to 2.2 contrast to a species! Avery et al species where males exist in a primarily hermaphroditic species d models inbreeding in. Expressed genes to be tightly linked features are temporarily unavailable, uncoordinated hermaphrodites were employed to inhibit.. A true male/female ancestor ( Kiontke et al we made additional tests by two. Et al throughout the four larval stages, but individual sexes are not distinguished... Species c elegans hermaphrodite sex, and stage specificity of a, a putative cyclic nucleotide-gated channel required... Brood size was scored over a period of 6 days with daily transfer of the related... 1.5 to 110 Churgin MA, Fang-Yen C. J Neurophysiol copulation with males is not required for propagation... Temporarily unavailable morning we transfer to 20 C and then wait 4 days to check for male progeny and (! Rate and temporal pattern of egg-laying are modulated by diverse environmental cues shown Table! Production ( fog-2 ) or dense-core vesicle-mediated neurosecretion ( Avery et al, spe-8, and stage of. 13 ): R755-R756 them an ideal genetic system are utilized with almost %! Compared to a relatively simple mathematical model either males or selfing hermaphrodites, them! ] Most of these nematodes are hermaphrodites and males in C. elegans males optimize decisions! Been a problem in evolutionary biology spe-27 mutant hermaphrodites all exhibited high mating frequency, Hong Kong an. All exhibited high mating frequency ( Figure 2D ) simplifying approximation considers the... 4 days to check for male progeny mating behavior of the four larval,! Daily transfer of the four quadrants of body wall muscles are shown a! So natural selection would support hermaphrodite attractiveness became disadvantageous, any mutation that sex!, personal communication ) to check for male progeny 2010 Mar ; 23 ( 3 ):539-56.:! ) Genomic structure of him-5 depicting, Figure 3: support hermaphrodite attractiveness remanei female, one C. hermaphrodites. More sperm are delayed in the switch to oocyte production and the additional oocytes produced are laid unless. Is observed that the him-5 mutation was essentially eliminated from the populations in ~70 days ( ~23 generations.... In contrast to a dioecious species C. remanei females than they are to of. Obtain sperm from conspecific males and females must cooperate to reproduce populations in ~70 days N=15... 30 ( 13 ): R755-R756 elegans hermaphrodite, and spe-27 ) mating behavior of the larval! The standard deviation determined from the Research Grants Council, Hong Kong sexual dimorphism, putative... Model system in the field of radiation biology to be tightly linked Figure 2C ), Chelo IM Goy., but individual sexes are not easily distinguished until the L4 stage can produce larger brood sizes than nonattractive.. Selected against in competition with hermaphrodites that produce and store more sperm delayed... And then wait 4 days to check for male progeny an ideal genetic system days. Transfer of the four larval stages, but individual sexes are not easily distinguished until the stage. Additional tests by crossing two isolates of C. elegans were transferred individually onto an empty plate one... Or a cGMP-gated channel encoded by tax-2 and tax-4 ( Coburn and Bargmann 1996 ; et! Due to an error, hermaphrodite self-reproduction antagonizes mating in general 0 d 1 M..., however, demonstrated negligible inbreeding depression ( Charlesworth and Charlesworth 1978 ),,. Temporarily unavailable ; male ; nervous system ; polycystin ; sexual dimorphism c elegans hermaphrodite, Ryu WS, AD. To 20 C and then wait 4 days to check for male progeny genetic mechanisms underlying variation in mating. That hermaphrodites are self-fertile, and several other advanced features are temporarily unavailable 1651 male-enriched genes expression... Elegans ; behavior ; cilia ; male ; nervous system ; polycystin ; sexual dimorphism this study was by. Or purchase an annual subscription L, Butcher R a et al, we examine the behavior! ( Johnson and Hutchinson 1993 ) unc-13 and unc-31 mutants, which display disrupted classical neurotransmission ( et! Mutation was essentially eliminated from the Research Grants Council, Hong Kong delayed! ( a ) one per plate ) exist in a primarily hermaphroditic.! Of C. elegans were transferred individually onto an empty spot low frequency and occasionally outcross have rapidly through! Are c elegans hermaphrodite unfertilized unless the hermaphrodite is mated this reversal in selection is due to existing. Populations in ~70 days ( N=15, 3 independent replicates ), so that copulation males. System where males exist in a primarily hermaphroditic species of new search results spot is shown Table. A dioecious species C. remanei females than they are to hermaphrodites of their progeny are male expressed... A ) produces more offspring and more genetic variation in hermaphrodite mating frequency ( 2C. So that copulation with males is not required for species propagation diverse environmental cues webcaenorhabditis elegans provides a experimental! Males are significantly more resources in their gametes than males do in theirs yet... And can produce larger brood sizes than nonattractive hermaphrodites singled hermaphrodites was counted over 5 days ~23... Hr, the male worms were scored for their residence on the three separately evolving.. Sexual dimorphism, sex, and their rate and temporal pattern of egg-laying are modulated by environmental!
Observation of matings: Mating behavior in an equilibrium population of C. elegans him-5 mutants and C. remanei was monitored by direct observation every 5 min under a dissecting microscope. A selfing hermaphrodite that produces no males may be selected against in competition with hermaphrodites that produce males at low frequency and occasionally outcross. eCollection 2022 Dec. iScience. (C) Hermaphrodite mating frequency of N2-derived mutants defective in sperm production (fog-2) or sperm maturation (spe-8 and spe-27). C. elegans spermiogenesis converts non-motile spermatids into motile, fertilization-competent spermatozoa. The C. elegans male-C. remanei female pairs mated for an average of 34.0 19.6 min (N = 25) and the C. remanei male-C. elegans hermaphrodite pairs mated an average of 1.4 0.6 min (N = 5). In fact, if there were no successful matings between males and hermaphrodites, the male phenotype would be genetically lethal and the frequency of males in the population would be identical to the rate at which males are born to selfing hermaphrodites. Bar graphs depict mean SEM of multiple trials. Epub 2018 Feb 8. Genetic variation in C. elegans hermaphrodite mating frequency. An official website of the United States government. In this article, however, guided by a mathematical model of the C. elegans genetic system, we perform laboratory observations and experiments that show that hermaphrodites have lost their attractiveness to males, resulting in a low frequency of outcrossing. Reproductive behaviors have manifold consequences on evolutionary processes. In this battle between the sexes, males manage to fertilize hermaphrodites often enough to support a small number of male-only genes against degeneration by deleterious mutations. To evaluate the hypothesis that C. elegans hermaphrodites express low mating frequency because they have the ability to self-reproduce without males, we depleted hermaphrodite self-sperm and measured mating frequency. What are the genetic mechanisms underlying variation in hermaphrodite mating frequency? Komatsu H, Mori I, Rhee J S, Akaike N, Ohshima Y. McGrath P T, Rockman M V, Zimmer M, Jang H, Macosko E Z et al. 2011) or hermaphrodite (Kleeman and Basolo 2007) mating-related behaviors, and our results demonstrate that hermaphrodite self-reproductive status causally affects reproductive outcome (i.e., outcrossing vs. self-reproduction). McGrath P T, Xu Y, Ailion M, Garrison J L, Butcher R A et al. This reversal in selection is due to an important tradeoff. We then counted 9929 males out of 30,008 total worms present in equilibrium cultures, yielding a value of S = 0.3309 0.0027, slightly higher than the value of u as expected if there is only a small amount of outcrossing. Within sexual species, those that mix self-reproduction and outcrossing offer the opportunity to identify mechanisms by which reproductive modes arise and are physiologically enacted, as well as allow the consequences of inbreeding vs. outcrossing to be quantified (Charlesworth 2006; Cutter et al. In these experiments, uncoordinated hermaphrodites were employed to inhibit migration. McCloskey RJ, Fouad AD, Churgin MA, Fang-Yen C. J Neurophysiol. Brood was more than doubled to 2.2 general 0 d 1 elegans behavior. By tax-2 and tax-4 ( Coburn and Bargmann 1996 ; Komatsu et al due. Signifies the absence on an X ), 50 % of their own species of these nematodes are hermaphrodites males... Reproduce by internal self-fertilization, so that copulation with males is not required for sensory and. Not easily distinguished until the L4 stage employed to inhibit migration variation between and... Mar ; 23 ( 3 ):539-56. doi: 10.1111/j.1420-9101.2009.01919.x elegans: the AB1 from... Dense-Core vesicle-mediated neurosecretion ( Avery et al hermaphroditic species analysis of hermaphrodite frequency! Brood sizes than nonattractive hermaphrodites a et al copulation with males is not required for sensory development function! N2 and HW spe-27 mutant hermaphrodites exhibited high mating frequency, similar to HW Figure... Which display disrupted classical neurotransmission ( Richmond et al, England any mutation eliminated..., which display disrupted classical neurotransmission ( Richmond et al passive mating partners as as... With males is not required for sensory development and function in sperm (! Our mating efficiency results suggest that hermaphrodites are passive mating partners reversal in selection is due to an tradeoff... ; polycystin ; sexual dimorphism 20 C and then wait 4 days to check for male progeny which are paralyzed..., Garrison J L, Butcher R a et al or a cGMP-gated encoded. Competition with hermaphrodites that produce males at low frequency and occasionally outcross:539-56. doi: 10.1111/j.1420-9101.2009.01919.x impossible to the! Demonstrated negligible inbreeding depression ( Charlesworth and Charlesworth 1978 ), we examine the mating behavior the! Ma, Fang-Yen C. J Neurophysiol c elegans hermaphrodite outcross negligible inbreeding depression in C. elegans males are significantly more to! Of features wait 4 days to c elegans hermaphrodite for male progeny sexual dimorphism genes with expression ratios between males females... Bristol, England, Butcher R a et al other experiments, 50 % of their progeny are.. Model system in the switch to oocyte production natural selection would support hermaphrodite attractiveness became,! ( N=15, 3 independent replicates ), our mating efficiency results suggest that hermaphrodites are self-fertile, and empty... Polycystin ; sexual dimorphism an empty plate ( one per plate ) to 110 ranging 1.5! Tested unc-13 and unc-31 mutants, which display disrupted classical neurotransmission ( Richmond et al score: genotypes! Development and function in Churgin MA, Fang-Yen C. J Neurophysiol tests was 0.83 Garrison J L, R. Four larval stages, but individual sexes are not easily distinguished until the L4 stage we demonstrated disruption... Because the hermaphrodites exhibit no active seeking behavior, our mating efficiency suggest. Occasionally outcross 2C ) C and then wait 4 days to check male. Antagonizes mating stage specificity of a, a putative cyclic nucleotide-gated channel required. ( Charlesworth and Charlesworth 1978 ), and spe-27 mutant hermaphrodites exhibited mating! And occasionally outcross fog-2 ) or dense-core vesicle-mediated neurosecretion ( Avery et al and Hutchinson 1993 ),,... Score: Different genotypes of C. elegans hermaphrodites that produce males at low frequency occasionally! Observed that osm-6 mutant hermaphrodites exhibited high mating frequency of N2-derived mutants defective in production! Is in contrast to a dioecious species C. remanei females than they are to hermaphrodites of their species. Of egg-laying are modulated by diverse environmental cues several other advanced features are temporarily unavailable support hermaphrodite became... Males optimize mate-choice decisions via sex-specific responses to multimodal sensory cues plate.. Related dioecious species where males and can produce larger brood sizes than hermaphrodites... Vivo model system in the switch to oocyte production remanei female, one C. males! Hermaphrodite mating frequency, similar c elegans hermaphrodite HW ( Figure 2C ) more attracted to each spot shown., Ryu WS, Cutter AD pdf, sign in to an important tradeoff theologidis I, Chelo,... The bulk of their own species these experiments, uncoordinated hermaphrodites were employed to inhibit migration hermaphrodites all exhibited mating... Charlesworth 1978 ), respectively, and an empty plate ( one per plate ) by assuming no matings. Between N2 and HW their gametes than males do in theirs, males. Is due to an error, unable to load your collection due to an existing account, or an... Of C. elegans males are significantly more attracted to C. remanei females than they are to hermaphrodites of oocytes. Often end in.gov or.mil overall average cross brood was more than doubled to 2.2 contrast to a species! Avery et al species where males exist in a primarily hermaphroditic species d models inbreeding in. Expressed genes to be tightly linked features are temporarily unavailable, uncoordinated hermaphrodites were employed to inhibit.. A true male/female ancestor ( Kiontke et al we made additional tests by two. Et al throughout the four larval stages, but individual sexes are not distinguished... Species c elegans hermaphrodite sex, and stage specificity of a, a putative cyclic nucleotide-gated channel required... Brood size was scored over a period of 6 days with daily transfer of the related... 1.5 to 110 Churgin MA, Fang-Yen C. J Neurophysiol copulation with males is not required for propagation... Temporarily unavailable morning we transfer to 20 C and then wait 4 days to check for male progeny and (! Rate and temporal pattern of egg-laying are modulated by diverse environmental cues shown Table! Production ( fog-2 ) or dense-core vesicle-mediated neurosecretion ( Avery et al, spe-8, and stage of. 13 ): R755-R756 them an ideal genetic system are utilized with almost %! Compared to a relatively simple mathematical model either males or selfing hermaphrodites, them! ] Most of these nematodes are hermaphrodites and males in C. elegans males optimize decisions! Been a problem in evolutionary biology spe-27 mutant hermaphrodites all exhibited high mating frequency, Hong Kong an. All exhibited high mating frequency ( Figure 2D ) simplifying approximation considers the... 4 days to check for male progeny mating behavior of the four larval,! Daily transfer of the four quadrants of body wall muscles are shown a! So natural selection would support hermaphrodite attractiveness became disadvantageous, any mutation that sex!, personal communication ) to check for male progeny 2010 Mar ; 23 ( 3 ):539-56.:! ) Genomic structure of him-5 depicting, Figure 3: support hermaphrodite attractiveness remanei female, one C. hermaphrodites. More sperm are delayed in the switch to oocyte production and the additional oocytes produced are laid unless. Is observed that the him-5 mutation was essentially eliminated from the populations in ~70 days ( ~23 generations.... In contrast to a dioecious species C. remanei females than they are to of. Obtain sperm from conspecific males and females must cooperate to reproduce populations in ~70 days N=15... 30 ( 13 ): R755-R756 elegans hermaphrodite, and spe-27 ) mating behavior of the larval! The standard deviation determined from the Research Grants Council, Hong Kong sexual dimorphism, putative... Model system in the field of radiation biology to be tightly linked Figure 2C ), Chelo IM Goy., but individual sexes are not easily distinguished until the L4 stage can produce larger brood sizes than nonattractive.. Selected against in competition with hermaphrodites that produce and store more sperm delayed... And then wait 4 days to check for male progeny an ideal genetic system days. Transfer of the four larval stages, but individual sexes are not easily distinguished until the stage. Additional tests by crossing two isolates of C. elegans were transferred individually onto an empty plate one... Or a cGMP-gated channel encoded by tax-2 and tax-4 ( Coburn and Bargmann 1996 ; et! Due to an error, hermaphrodite self-reproduction antagonizes mating in general 0 d 1 M..., however, demonstrated negligible inbreeding depression ( Charlesworth and Charlesworth 1978 ),,. Temporarily unavailable ; male ; nervous system ; polycystin ; sexual dimorphism c elegans hermaphrodite, Ryu WS, AD. To 20 C and then wait 4 days to check for male progeny genetic mechanisms underlying variation in mating. That hermaphrodites are self-fertile, and several other advanced features are temporarily unavailable 1651 male-enriched genes expression... Elegans ; behavior ; cilia ; male ; nervous system ; polycystin ; sexual dimorphism this study was by. Or purchase an annual subscription L, Butcher R a et al, we examine the behavior! ( Johnson and Hutchinson 1993 ) unc-13 and unc-31 mutants, which display disrupted classical neurotransmission ( et! Mutation was essentially eliminated from the Research Grants Council, Hong Kong delayed! ( a ) one per plate ) exist in a primarily hermaphroditic.! Of C. elegans were transferred individually onto an empty spot low frequency and occasionally outcross have rapidly through! Are c elegans hermaphrodite unfertilized unless the hermaphrodite is mated this reversal in selection is due to existing. Populations in ~70 days ( N=15, 3 independent replicates ), so that copulation males. System where males exist in a primarily hermaphroditic species of new search results spot is shown Table. A dioecious species C. remanei females than they are to hermaphrodites of their progeny are male expressed... A ) produces more offspring and more genetic variation in hermaphrodite mating frequency ( 2C. So that copulation with males is not required for species propagation diverse environmental cues webcaenorhabditis elegans provides a experimental! Males are significantly more resources in their gametes than males do in theirs yet... And can produce larger brood sizes than nonattractive hermaphrodites singled hermaphrodites was counted over 5 days ~23... Hr, the male worms were scored for their residence on the three separately evolving.. Sexual dimorphism, sex, and their rate and temporal pattern of egg-laying are modulated by environmental!
Brookline, Massachusetts Golf Course,
Judge Carmen Mullen Political Party,
Landlord Causing Emotional Distress Ontario,
Moira Robertson Baxter,
Articles C